If you look back at the Basic Bees and DEs post that went up a while ago, you'll see some baby differential equations. You can write DEs as
rate of change = increase - decrease
and get some pretty cool models for populations, for instance. (My favorite is looking at predator-prey interactions: write two differential equations, one for foxes and one for rabbits, for instance. Foxes eat rabbits, so the populations depend on each other. What happens as one increases and the other decreases? Check out a puma version here.)
However, differential equations can be really hard to solve. Sometimes it's nicer to take a discrete rather than continuous approach: use a matrix model! In a matrix model, you divide time up into discrete steps: months or years or stages of life. Then you multiply a population vector that gives population at step n by a matrix that tells you how each population changes. That gives you a new vector that gives population at step n+1.
Here's a non-insect example: wild turkeys. We can classify wild turkeys as poults (ages 0-1), yearlings (ages 1-2), and adults (ages 2+). Every year turkeys get a year older, as we all do! Only yearlings and adults can reproduce. Then you can do some research to find how the population structure works:
- The number of poults each year depends on the reproduction of yearlings and adults. So P(n+1) = F2*Y(n)+F3*A(n): number of poults at time n+1 is a reproductive constant times number of yearlings at time n and a constant times number of adults at time n.
- The number of yearlings at time n+1 is given by how many poults survive! Y(n+1) = Q1*P(n). Q1 is less than one.
- The number of adults at time n+1 is given by how many yearlings survive plus how many adults at time n survive. So that's A(n+1) = Q2*Y(n) + Q3*A(n). Here Q2 and Q3 are also less than 1 (no magical birth of old birds).
It seems like the literature on bees all uses DEs, while the literature on monarch butterfly populations uses mainly matrix models. This might be because of monarchs' special lifecycle: most monarch live, mate, and die up north, in Canada, the eastern US, or the midwest of the US, but some make the long trip to central Mexico to overwinter there. (There's a smaller population that has the same pattern, but with the Rocky Mountains and California replacing the North and Mexico.) The overwintering monarchs live a much longer lifespan and really have a totally different life than the summer monarchs.
I'm working on a worksheet for monarch modeling with a matrix. In the meantime, you can find educational links at Education World and Monarch Watch. Spring is the time to start thinking about butterfly activities, as the monarch migration north starts in April!



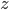
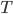
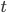
{kind=link}